 |
| Palm fruits are mostly single-seeded drupes, brightly colored to attract birds or other animals for dispersal. The large, hard, seeds either pass unharmed through the digestive system or are dropped to the ground as the fleshy pericarp is eaten. |
Palm fruits may seem like an odd place to look for sclerenchyma, but I discovered early in my career as a plant taxonomist specializing in palms, that not only are such cell types present, but they are also highly varied in type and arrangement. They represent an excellent case study for the usefulness of comparative plant anatomy.
One of the functions of sclerenchyma in general is to protect plant tissues from vegetation-chomping animals, and fruits are one of the most vulnerable of plant organs. Fruits, and the seeds within, fill up with valuable nutrients as they mature. The seeds must obviously be protected until they can be dispersed and have a chance to produce the next generation. Fruits, however, are often meant to be eaten as part of that dispersal, but not until the seeds are mature. So unripe fruits must be protected until then, but must become palatable, sometimes quickly and dramatically, at maturity.
The first layer of defense for the large seed within a palm fruit is something called the locular epidermis. This is actually the interior epidermis of the carpel that surrounds the seed proper. In this layer, the cells often elongate perpendicular to the fruit wall, become pillar-like, closely-packed sclereids as the fruit matures. Similar layers of cells have independently evolved in the seed coats of legumes.
A well-developed locular epidermis is common among palms of the subtribe Areceae (the large, advanced group that includes the betel nut, Areca catechu), but is quite varied in thickness, even within genera. Where it is not present, other forms of sclerenchyma take its place.
Another type of sclerenchyma found in palm fruits consists of individual cells resembling grains of sand, called brachysclereids or stone cells. Those found in palms are similar to the gritty patches of stone cells found just below the epidermis in pear fruits. Stone cells may be scattered within parenchyma tissue, grouped in clusters, or found in continuous layers.
In a great many palm fruits, there are also many fibrous bundles, consisting of narrow, thick-walled fiber cells. As is generally true in vascular plants, fibers occur mostly around strands of vascular tissue (xylem and phloem), as protection for those tissues. When additional protective functions (as in palm fruits) or supportive functions (i.e. in wood or the fibrous stems of palms and bamboos) are present, the volume of fibrous tissue can become massive and far in excess of what is needed to protect the vascular tissues.
 |
| The fruits of Rhopaloblase ceramica have a very thick locular epidermis (bottom layer), consisting of elongate, pillar-like sclereids, packed tightly together. Above that, are three tiers of massive fibrous bundles that form around vascular tissues. In a band below the outer epidermis, are scattered stone cells (brachysclerieds), stained a purplish red. The very dark tissues present contain tannins. |
Often intermixed with the fibrous vascular bundles close to the seed is a tissue with the seemingly oxymoronic name of sclerified parenchyma. This is a region that begins as normal parenchyma in the young fruit, but become "sclerified" (develop secondary walls) as the fruit reaches its full size. In some of my earlier papers, I referred to this as "sclerified ground tissue," but that was too vague, as there are other forms of sclerenchyma in the ground tissue. (Ground tissue refers to the tissue that fills the interior of leaves, stems, roots, and fruits, and consists mostly of parenchyma.
In the most specialized of the bird-dispersed palm fruits, we can usually see three distinct zones:
Close to the seed, we find densely packed fibrous bundles, sclerified parenchyma, and often a thickened locular epidermis. These hard tissues are typically fused together into a solid endocarp, or pit, which remains with the seed when the rest of the fruit is removed. This helps prevent the crushing of the seed when the fruit is eaten, or its penetration by burrowing insects.
Below the outer epidermis, one can often see an exocarp, a layer of stone cells, cells filled with bitter tannins, and sometimes fibrous bundles that protect against insect penetration. Stone cells, which occur individually or in small patches, are particularly advantageous in this outer fruit region because they can loosen and separate as the fruit expands. This allows the fruit to swell as it ripens, becoming more succulent.
Between these outer and inner protective layers, is the larger expanse of tissue referred to as the mesocarp. This middle region may also be filled with fibrous bundles (the fibrous outer husk of the coconut being an extreme example), but in many specialized fruits it has been cleared of hard tissues, and consists only of soft parenchyma, which can swell as the fruit ripens, becoming fleshy, tasty and nutritious. Such fruits presumably provide the most food for birds that feed upon them, and so have a selective advantage.
Such well-defined zones are particularly conspicuous in the Ptychosperma alliance, which I studied as a graduate student. In this group of palms native to New Guinea, Australia and some Pacific islands, another extraordinary transformation has taken place: the evolution of two radically different kinds of fibrous bundles, one occupying the endocarp, the other occupying the exocarp.
 |
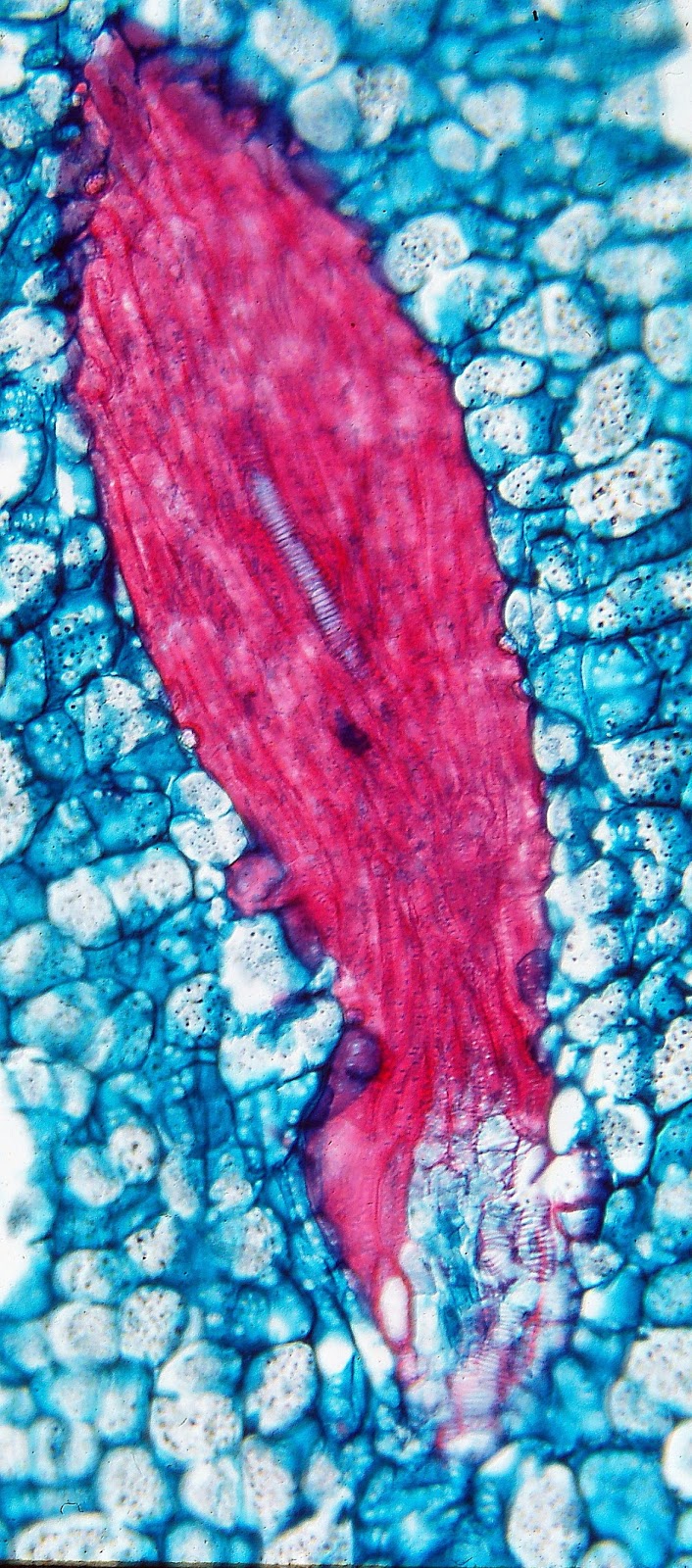
| In Heterospathe, "naked" bundles of vascular tissue are at the bottom, close to the seed, while bundles further out contain only fibers. |
 |
| In the short spurs of fibers in Orania, bits of vascular tissue (ladder-like protoxylem element in center) can be found, illustrating the role procambia in forming fibrous bundles. |
 |
 In Veitchia, inner bundles contain small strands of vascular tissue (white spots) and thick fibrous sheaths. Bundles in the outer half of the fruit are purely fibrous. Legend applies to all the diagrams. |
 |
| In Veitchia, the outer fibrous bundles are elongate and parallel with the surface, but can separate from one another as the fruit expands. |
In the least specialized members of the Ptychosperma alliance, such Veitchia and Normanbya, fibrous bundles have already been separated into two distinct groups, the inner bundles have at least some vascular tissue and form an interconnected network, while the outer bundles are devoid of vascular tissue altogether, and become disconnected from one another as the fruit expands.
In the remaining genera of the Ptychosperma alliance, the outer fibrous bundles have become quite short and clearly separate from one another. They are confined to the exocarp and are mixed with the brachysclereids. Variation on the arrangement of tissues, however, is significant, and can be used to identify the different genera. Some examples are below, but so that this post won't get too long, I refer you to my original paper on the Ptychosperma alliance, for more details. Similar trends can be seen in the other alliances of the subtribe Areceae, also with distinctive arrangements in different genera, and papers on those can be accessed through my general list of publications.
 |
| In Ptychosperma and other advanced genera, inner fruit tissues follow the distinctive grooves of the seed. The outer fibrous bundles are small, short, and perpendicular to the surface. |
 |
| In Ptychosperma, outer fibrous bundles are short and perpendicular to the surface. Brachysclereids fill in between them. |
 |
| An isolated outer fibrous bundle from Brassiophoenix. |
 |
| The large fruits of Ptychococcus have an exceptionally thick, hard endocarp, consisting of a thick locular epidermis, a massive layer of sclerified parenchyma, and a mantle of fibers formed from the fusion of adjacent fibrous vascular bundles. Short fibrous bundles also mingle with brachysclereids in the exocarp. |
 |
| The fruits of Brassiophoenix have an angular endocarp, like that of Ptychococcus, but with the fibrous vascular bundles embedded within the sclerified parenchyma. |
No comments:
Post a Comment
Note: Only a member of this blog may post a comment.